Phosphatase Subfamily DSP1
Phosphatase Classification: Superfamily CC1: Family DSP: Subfamily DSP1
DSP1 is an inducible nuclear MAP Kinase phosphatase (MKP) conserved throughout eukaryotes. As a key player in MAPK pathway, it is implicated in immune regulation and cancer.
Evolution
DSP1 subfamily is widely found across eukaryotes. It is present in animals, plants, amoeba, and some basal eukaryotes, but is absent from ecdysozoa (nematodes and arthropods), most fungi and Monosiga (unpublished data, DUSP1, DUSP2, DUSP4, DUSP5).
Domain Structure
DSP1 has an inactive rhodanese domain, followed by a phosphatase domain. In general, a kinase interaction motif (KIM) embedded in rhodanese domain mediates the interaction between DSP1 and MAPK (i.e. ERK, JNK, p38 kinase subfamilies) [1, 2, 3, 4].
Dictyostelium has four DSP1s. Two (mkpB-1 and mkpB-2) have the typical domain combination of rhodanese domain and phosphatase domain. Another two have different domain combination. Both of them have a long N-terminal region. One of the two, mkpA, has three weak hits to Gelsolin domain, an actin-binding protein that is a key regulator of actin filament assembly and disassembly.
Functions
In general, DSP1 are inducible nuclear MAP Kinase phosphatases (MKPs). Given the MAPKs modules many different biological processes, it is not surprising that DSP1 subfamily functions in cancer and immune system. Human members are DUSP1, DUSP2, DUSP4 and DUSP5. Their substrate specificity, subcellular localization and etc have been summarized in Table 1 in review paper [5]. Although they have overlapping substrate specificity, there are differences in their mRNA regulation, response to extracellular stimuli, and tissue-specific expression, suggesting they serve specific roles in cellular function [6].
DUSP1 (MKP1/hVH1)
DUSP1 is the best characterized phosphatase within this subfamily. It prefers to dephosphorylates p38 and JNK over ERK. DUSP1 is involved in immune regulation and cancer. DUSP1 is an important negative-feedback regulator of macrophage function (possibly by modulating mRNA-destabilizing protein tristetraprolin [7]) and the inflammatory response to TLR signalling, and plays key regulatory roles in both innate and adaptive immune responses via inactivation of p38 and JNK (reviewed in [5]). DUSP1 is an important negative regulator of pro-allergic responses in airway epithelium [8].
DUSP1 (MKP1) also mediates G1-specific dephosphorylation of H3Serine10P in histone in response to DNA damage. The phosphorylation at H3S10 plays a dual role in a cell by maintaining relaxed chromatin for active transcription in interphase and condensed chromatin state in mitosis [9].
(Note: It is worthy pointing out that DUSP1 expression is strictly regulated at low level in brain (see GTEx)).
DUSP2 (PAC1)
As a MKP, DUSP2 has preference toward ERK=p38 > JNK. DUSP2 is inactive when alone in vitro, and dephosphorylates extracellular signal-regulated kinase 2 (ERK2) but not p38alpha or c-Jun NH(2)-terminal kinase 2 (JNK2). ERK2 dephosphorylation by DUSP2 requires association of its rhodanese domain and ERK2 that results in catalytic activation of the phosphatase. p38alpha interacts with but does not activate DUSP2, whereas JNK2 does not bind to or cause catalytic activation by DUSP2 [2, 3].
It has been shown that individual mutation of the conserved Arg294 and Arg295 that likely comprise the phosphothreonine-binding pocket in DUSP2 to either alanine or lysine results in a nearly complete loss of its phosphatase activity even in the presence of ERK2 [3]. (PS: However, the conservation is not found within DSP1 subfamily. On the other hand, they are found in other subfamilies no matters they dephosphorylate ERK or not, which suggested the two arginine residues do not determine phosphatase activity or the specific phosphatase activity towards ERK.)
DUSP2 is expressed in various tissues (see GTEx). Its expression is regulated by p53 which binds to palindromic site in DUSP2 promoter [10]. Its expression is regulated, or more precisely speaking, inhibited by hypoxia inducible factor-1 (HIF-1) [11, 12].
DUSP4 (MKP2/TYP)
DUSP4 prefers ERK and JNK over p38 [13, 14, 15]. Further studies has shown the catalytic activity of DUSP4 was enhanced dramatically by ERK and JNK but was affected only minimally by p38. By contrast, p38 and ERK bound DUSP4 with comparably strong affinities, whereas JNK and DUSP2 interacted very weakly [16]. DUSP4's substrate in vivo is controversial. As reported in [17], DUSP4 selectively dephosphorylates JNK, or different DUSP4 isoforms have different substrate specificity as shown in [18].
DUSP4 dephosphorylates ERK, and on the other hand, ERK can phosphorylates DUSP4 (on Ser386 and Ser391 at its C-terminus) which leads to stabilization of DUSP4 protein, as blockage of ERK activation results in enhanced proteasomal degradation of DUSP4 protein [19]. (Note: Ser386 and Ser391 are also found in DUSP1, which preferential dephosphorylates p38 and JNK but NOT ERK. DUSP2 has Ser386 but position 391 is Ala; DUSP5 does not align at C-terminal region.)
DUSP4 is involved in cancer. For instance, it is a common epigenetically silenced gene in glioma [20]. DUSP4 is frequently upregulated in breast malignancy [21].
DUSP4 is also involved in inflammation. It is a negative regulator of macrophage M1 activation through JNK and p38 and inhibits inflammation during macrophage-adipocyte interaction [22].
(Note: look into the conservation of basic motif mentioned in [16, 23], which is supposed to mediate the interaction between DUSP4 (or even other DSP1 or MKPs) with ERK (or MAPKs in general).
DUSP5 (hVH3)
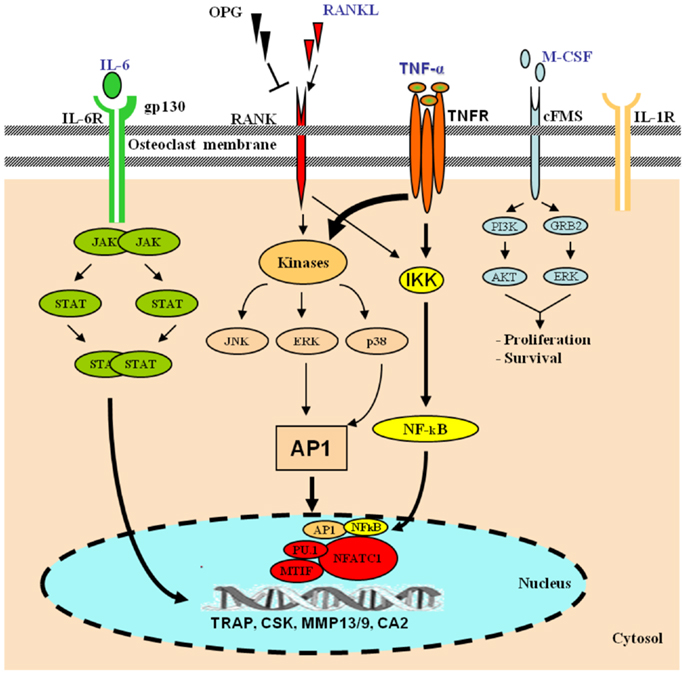
DUSP5 is an inducible nuclear ERK-specific MKP that functions as both an inactivator of and a nuclear anchor for ERK2 in mammalian cells [1, 24]. Thus, it modules T-cell development and function [25], modules corneal epithelial cell proliferation [26], is implicated in angiogenesis [27], and influences myeloid cell fate by M-CSF pathway [28]. It may be involved in carcinogenesis, since DUSP5 is down-regulated in gastric cancer by promoter CpG island hypermethylation [29], up-regulated in MCF-7 human breast carcinoma cells by transcription factor c-Jun [30] and its down-regulation predicts poor prognosis of patients with prostate cancer [31].
{kind=link}
Similar with DUSP4, DUSP5 is also stabilized by binding to and be phosphorylated by ERK. The accumulation of DUSP5 protein is regulated by rapid proteasomal degradation. DUSP5 is phosphorylated by ERK1/2 both in vitro and in vivo on three sites (Thr321, Ser346 and Ser376) at C-terminus. Co-expression of ERK2 results in significant stabilisation of DUSP5, which is accompanied by reduced levels of DUSP5 ubiquitination. However, there is no evidence that phosphorylation of DUSP5 by ERK2 significantly affects either the half-life of the DUSP5 protein or its ability to bind to, inactivate or anchor ERK2 in the nucleus. These changes are independent of ERK2 kinase activity but absolutely depend on the ability of ERK2 to bind to DUSP5 [4].
DUSP5 is regulated by p53 binding to DUSP5 promoter [32] (same mechanism as DUSP2).
References
Error fetching PMID 19843478:
Error fetching PMID 26064329:
Error fetching PMID 11387337:
Error fetching PMID 8626452:
Error fetching PMID 24308939:
Error fetching PMID 12575935:
Error fetching PMID 25638726:
Error fetching PMID 19801501:
Error fetching PMID 7535768:
Error fetching PMID 7961985:
Error fetching PMID 25816341:
Error fetching PMID 25834604:
Error fetching PMID 8545112:
Error fetching PMID 18430737:
Error fetching PMID 19666109:
Error fetching PMID 7836374:
Error fetching PMID 15713638:
Error fetching PMID 20554528:
Error fetching PMID 21490398:
Error fetching PMID 19228121:
Error fetching PMID 21084841:
Error fetching PMID 26111828:
Error fetching PMID 23402999:
Error fetching PMID 26019272:
Error fetching PMID 10655591:
Error fetching PMID 12944906:
Error fetching PMID 20124482:
Error fetching PMID 20806045:
Error fetching PMID 21984126:
Error fetching PMID 12673251:
Error fetching PMID 16288922:
- Error fetching PMID 15713638:
- Error fetching PMID 12575935:
- Error fetching PMID 16288922:
- Error fetching PMID 19666109:
- Error fetching PMID 19228121:
- Error fetching PMID 7836374:
- Error fetching PMID 26019272:
- Error fetching PMID 25638726:
- Error fetching PMID 26111828:
- Error fetching PMID 12673251:
- Error fetching PMID 21490398:
- Error fetching PMID 21984126:
- Error fetching PMID 7535768:
- Error fetching PMID 8545112:
- Error fetching PMID 8626452:
- Error fetching PMID 11387337:
- Error fetching PMID 16038800:
- Error fetching PMID 19843478:
- Error fetching PMID 21084841:
- Error fetching PMID 20124482:
- Error fetching PMID 25834604:
- Error fetching PMID 25816341:
- Error fetching PMID 10655591:
- Error fetching PMID 7961985:
- Error fetching PMID 18430737:
- Error fetching PMID 20806045:
- Error fetching PMID 24308939:
- Error fetching PMID 19801501:
- Error fetching PMID 23402999:
- Error fetching PMID 20554528:
- Error fetching PMID 26064329:
- Error fetching PMID 12944906: